
Techniques
 Optogenetics.
Optogenetics.Genetic or viral approaches are used to express light‑activated molecules in specific cells (e.g. channelrhodopsin or archaerhodopsin) to define the role of these neurons in circuits that mediate behavior and endocrine responses. Such experiments are performed in vivo or in vitro, where action potential firing is precisely suppressed by a pulse of laser light. Here, a neuron expressing the inhibitory opsin Archaerhodopsin is inhibited by the brief application of yellow light.
We perform in vivo and in vitro recordings from large populations of neurons using Neuropixel probes, including single and quad-shank probes.
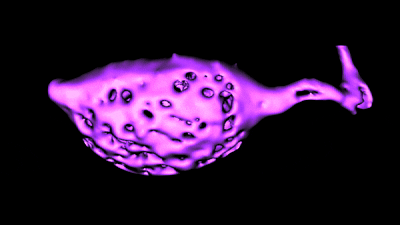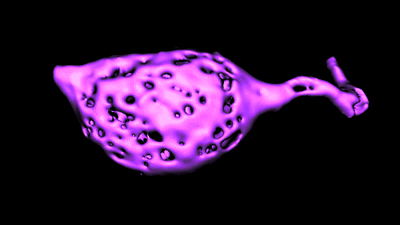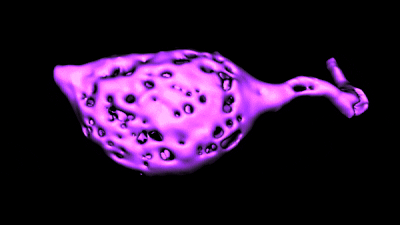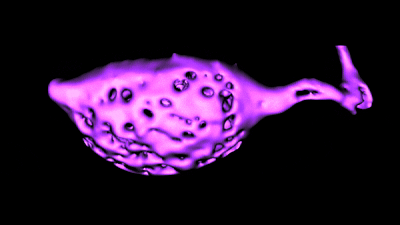 Histology and morphometry.
Histology and morphometry.We use immunohistochemistry, phalloidin staining for actin filaments, antibody staining of live or fixed cells, intracellular dye injections, and morphometric analysis of the surface area of single cells or neurons imaged in live tissue slices. We also use Diolistic labeling and various methods for tracing axonal pathways.
 Digital imaging.
Digital imaging.We perform dynamic analysis of changes in cell volume, ion concentration imaging (e.g. calcium), and fluorescence detection combined with electrophysiology.
Tissue culture and molecular biology.
We use organotypic slice cultures, cultured cell lines, in‑vitro transfection, western blotting, and RT‑PCR for detection of specific mRNA species in single cells and tissue extracts.
We use organotypic slice cultures, cultured cell lines, in‑vitro transfection, western blotting, and RT‑PCR for detection of specific mRNA species in single cells and tissue extracts.
 Extracellular single‑unit recording.
Extracellular single‑unit recording.A glass pipette filled with saline is used to record action potentials fired by a neuron at the tip of the pipette. This non‑invasive technique allows long‑duration recordings of spontaneous activity.
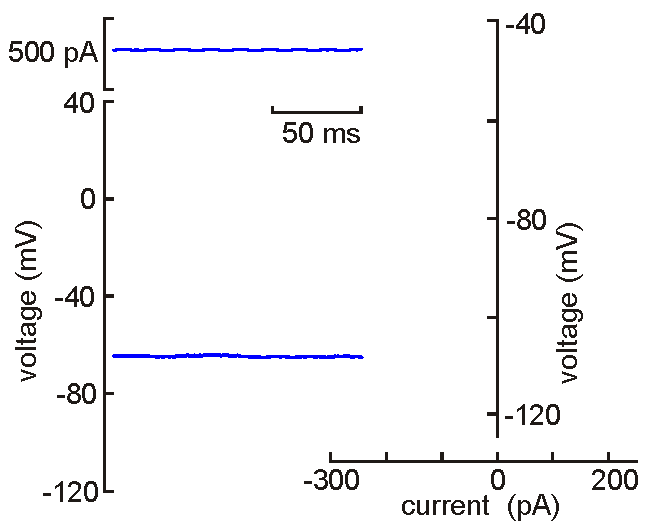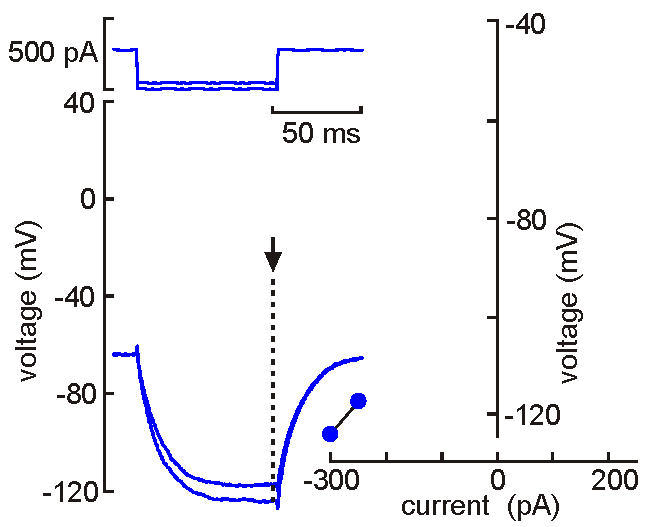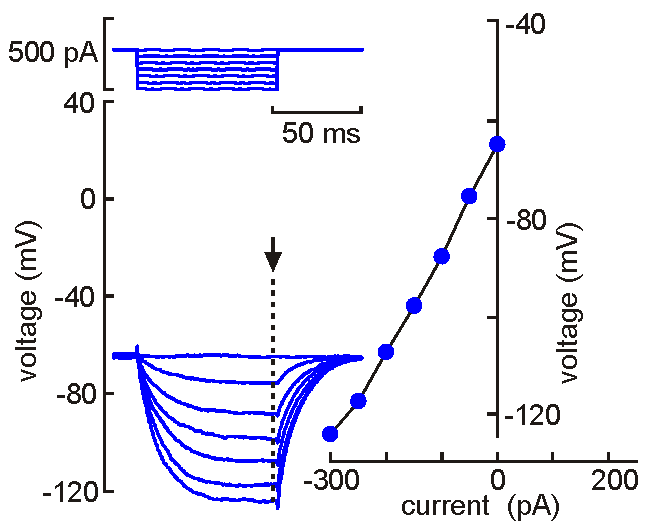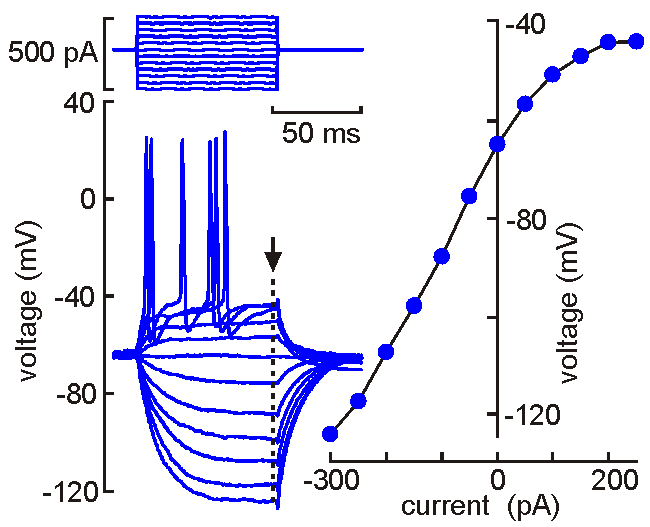 Intracellular (sharps) recording.
Intracellular (sharps) recording.A fine‑tipped micropipette (~100 nm) is inserted into the neuron, allowing direct recording of membrane potential, resistance, time constant, synaptic potentials, and action potentials. Voltage‑current analysis provides measures of input resistance.
 Patch clamp – single channel recording.
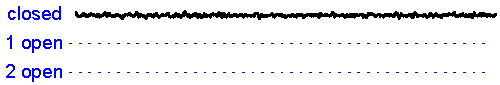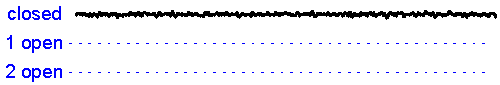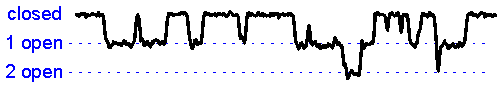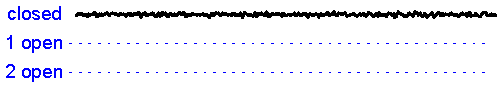
Patch clamp – single channel recording.A gigaseal allows recording of individual ion channel openings. Multiple channels can be observed simultaneously when the current amplitude doubles.
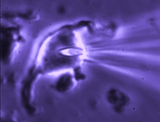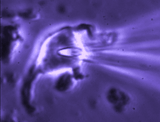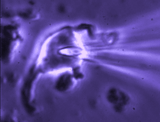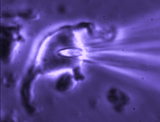
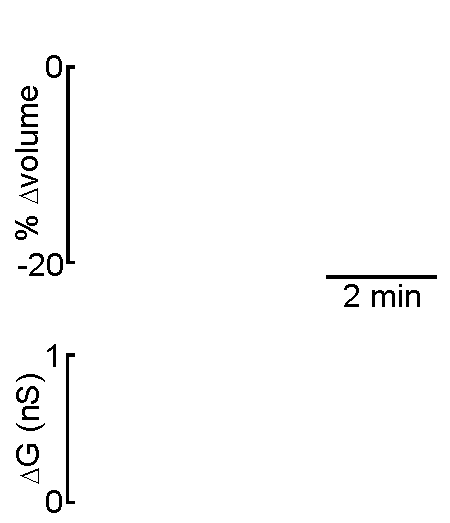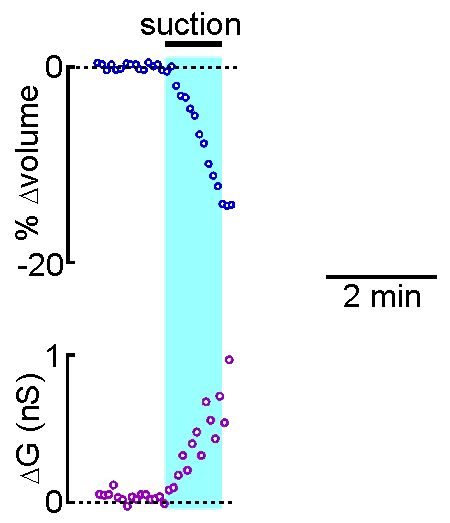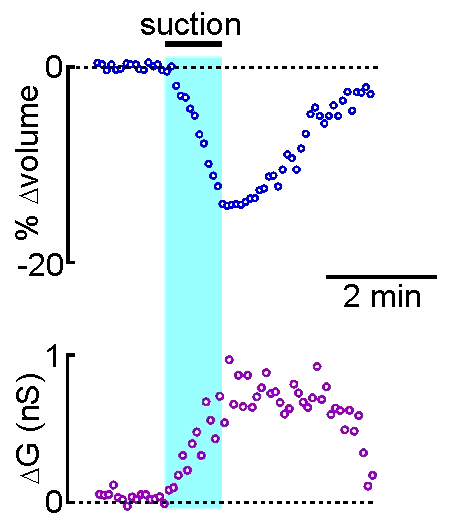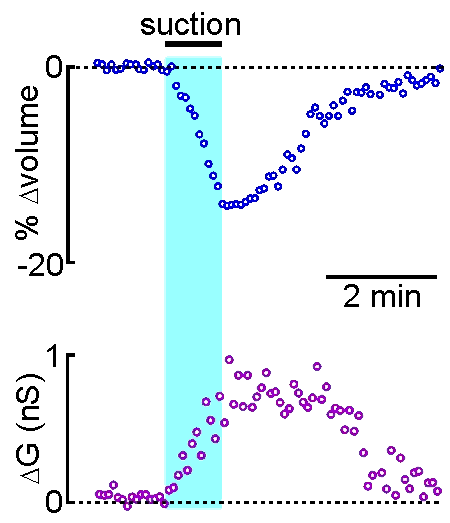 Patch clamp – whole cell recording.
Patch clamp – whole cell recording.After forming a gigaseal, a brief pulse ruptures the membrane patch, allowing direct electrical access to the cell interior. This enables current‑clamp and voltage‑clamp recordings, and delivery of dyes or fluorescent probes. Shrinking a neuron by suction increases membrane conductance, revealing mechanisms of osmosensory transduction.
 Fast drug application.
Fast drug application.Whole‑cell recordings are combined with rapid solution exchange using a piezo‑driven stepper system to study fast ion channel responses and synaptic activation.
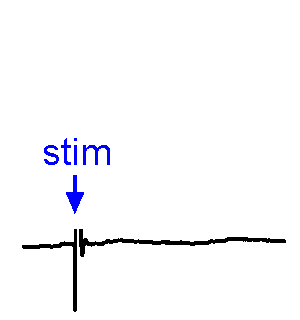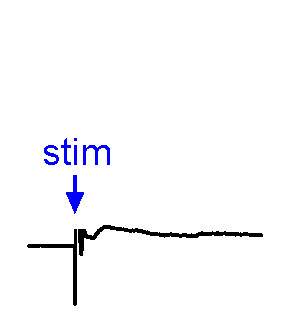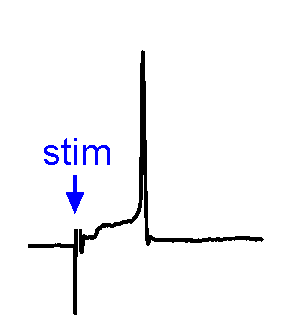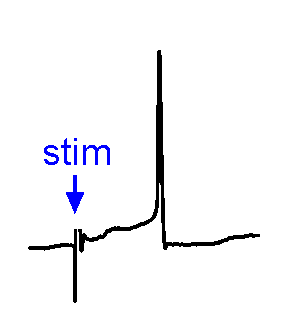 Synaptic responses.
Synaptic responses.We analyze spontaneous and evoked synaptic events. EPSPs may or may not trigger action potentials. We also study physiological stimulation of afferents, such as osmotic or thermal activation of sensory neurons. See the Research page for examples.
 Cells and tissues.
Cells and tissues.We study isolated somata, nerve terminals, acute slices, superfused explants, and organotypic cultures to examine neuron‑glia interactions and the integration of cellular, synaptic, and network properties.